EXPRESSION DE L'INFORMATION GENETIQUE
STRUCTURE DE L'INFORMATION GENETIQUE:
L'information génétique est définie par l'activité de la molécule d'ADN. L'ADN, support de l'information génétique, est formé par un arrangement caractéristique de l'espèce de quatre types de nucléotides qui se distinguent par leur bases azotées. Les nucléotides se reconnaissent deux à deux (A reconnaît T et réciproquement et C reconnaît G et réciproquement) ce qui fait que cette molécule est formée de deux chaînes complémentaires. Dans l'espace, cette molécule se présente en double hélice. Au cours de la multiplication de la molécule d'ADN, chaque chaîne ancienne construit une nouvelle chaîne qui lui est complémentaire et qui est identique à celle avec laquelle elle était associée. On parle, alors, d'une duplication semi-conservative de la molécule d'ADN.
EXPRESSION DE L'INFORMATION GENETIQUE:
Chaque individu présente un ensemble de caractères qui correspondent à son phénotype. Ces caractères sont le produit de l'expression du matériel génétique puisqu'ils sont transmissibles à travers les générations. L'expression de ces caractères revient à la présence de protéines spécifiques (c'est -à- dire à un arrangement spécifique en acides aminés). Plusieurs expériences telles que l'utilisation d'acides aminés radioactifs ont montré que la synthèse des protéines se fait dans le cytoplasme alors que l'information génétique est localisée dans le noyau. Ces observations ont laissé comprendre que la synthèse protéique ne se fait pas directement à partir de la molécule d'ADN d'où la nécessité d'un intermédiaire capable de se déplacer entre le noyau et le cytoplasme pour transporter l'information génétique jusqu'à la machine de synthèse protéique. Cet intermédiaire est l'ARN messager. Dans nos cellules il y a trois types d'ARN à savoir:
ARN messager noté ARNm.
ARN de transfert noté ARNt qui fait reconnaître les acides aminés à l'ARNm pour synthétiser les protéines.
ARN ribosomique noté ARNr qui rentre dans la structure du ribosome qui forme la machine de synthèse protéique.
Ainsi, on comprend que l'expression de l'information génétique se fait en deux étapes: on appelle transcription la conversion du gène en ARNm et on appelle traduction la conversion de l'ARNm en protéine c'est à dire en une séquence d'acides aminés.
LA TRANSCRIPTION:
La molécule d'ADN est formée de deux brins d'où elle est dite bi-caténaire alors que la molécule d'ARNm est mono-caténaire ce qui laisse comprendre que la synthèse de l'ARNm se fait à partir de l'un des deux brins de l'ADN qui sera appelé brin transcrit. Cette synthèse se fait selon la règle de complémentarité suivante:

Puisque l'ARNm synthétisé correspond à un gène, la synthèse doit commencer à partir d'un point précis appelé signal d'initiation pour se terminer au niveau d'un autre point précis appelé signal de terminaison. L'enzyme qui contrôle cette synthèse est appelé ARN polymérase. Ce mécanisme de la transcription est universel. Cependant, et comme le gène procaryote n'a pas la même structure que le gène eucaryote, l'ARNm synthétisé va être relativement différent.
Cas du gène procaryote:
L'observation du gène procaryote laisse constater que l'ARNm synthétisé est directement utilisé pour la synthèse protéique. Donc, chez les procaryotes, l'ARNm correspond nucléotide par nucléotide à la portion transcrite du gène.

Cas des eucaryotes:
Dans ce cas le gène est formé de deux catégories de séquences nucléotidiques:
Les exons:qui correspondent aux séquences codentes et se trouvent sous forme complémentaire dans l'ARNm exporté vers le cytoplasme.
Les introns: qui correspondent à des séquences non codentes, insérées entre les éxons et qui n'ont pas de complémentaires dans l'ARNm utilisé pour la synthèse protéique.
Le gène procaryote est, ainsi, dit morcelé ou en mosaïque. Suite à cette structure, la transcription du gène eucaryote se fait en deux phases: d'abord le gène est transcrit en sa totalité (exons et introns) pour donner un premier ARN dit ARN préméssager . Puis, au niveau même du noyau, l'ARN préméssager subit une excision des parties correspondantes aux introns et un épissage des parties correspondantes aux exons pour donner, en fin, l'ARNm qui sera transféré vers le cytoplasme.
LA TRADUCTION:
La traduction consiste à faire correspondre la séquence nucléotidique de l'ARNm à la séquence d'acides aminés de la protéine. La question qui s'impose est : comment peut-on faire correspondre les quatre nucléotides de l'information génétique aux vingt acides aminés des protéines?
Le code génétique:
La réponse à cette question a permis de définir le code génétique. Ce code a été mis en place à la suite d'un raisonnement mathématique qui a été confirmé par l'expérimentation. En effet, si on fait correspondre à un nucléotide un acide aminé (41=4) on n'obtient pas un arrangement satisfaisant puisqu'une protéine ne renfermera au maximum que quatre acides aminés différents! Ce qui n'est pas le cas dans la nature. De même, un arrangement qui fait correspondre deux nucléotides à un acide aminé n'est également pas satisfaisant (42=16). Seul, l'arrangement qui fait correspondre trois nucléotides à un acide aminé répond à la question puisqu'il donne soixante quatre combinaisons possibles ce qui couvre largement les vingt acides aminés naturels. Le triplet qui définit un acide aminé est appelé un codon d'où le code génétique est formé de soixante quatre codons. Ce code possède quatre propriétés:
Le code génétique est dégénéré: c'est-à-dire que la plupart des acides aminés sont définis par plus d'un seul codon.
Le code génétique n'est pas chevauchant: c'est à dire qu'un nucléotide n'appartient qu'à un seul codon et la lecture se fait codon par codon.
Le code génétique est à quelques exceptions près universel.
Trois codons ne définissent aucun acide aminé et sont ainsi appelés codons stop ou codons non sens. Il s'agit des codons UAA, UAG et UGA.
La machine de synthèse:
La machine de synthèse est formée d' acides aminés, de l'ARNm, de ribosomes (voir schéma) et de l'ARNt qui est formé d'une chaîne nucléotidique associée à des protéines et qui présente la forme d'une feuille de trèfle avec quatre pôles dont un qui forme un site de fixation de l'acide aminé et un autre qui est formé d'un triplet complémentaire au codon correspondant à l'acide aminé en question et qu'on appelle anti-codon.
Mécanisme de la traduction:
La traduction de l'ARNm en protéine se fait en trois phases successives:
La phase d'initiation:
L'ARNm, libre dans le cytoplasme, sera fixé par une petite sous-unité du ribosome tel qu'un codon initiateur (toujours le même) AUG sera exposé en premier plan ce qui va faire appel à une grande sous-unité du ribosome qui couvre l'ensemble. Cette sous-unité est formée de deux sites: site A et site P. Le site P se trouve déjà en face du codon initiateur ce qui fait appel à un ARNt initiateur puisqu'il porte l'anti-codon UAC et un acide aminé initiateur: la méthionine. La fixation de cet ensemble dans le site P déclenche la synthèse protéique.
La phase d'élongation:
Le site A de la grande sous-unité étant libre devant le deuxième codon, ceci fait appel à un deuxième ARNt qui porte le deuxième acide aminé. L'installation de cet ARNt dans le site A stimule la formation de la liaison peptidique entre les deux acides aminés et la rupture de la liaison entre le premier ARNt et son acide aminé ce qui rend le site P libre d'où le glissement du ribosome d'un codon. Ainsi, le site A devient libre devant le troisième codon et le site P héberge le deuxième ARNt chargé des deux premiers acides aminés ce qui stimule l'arrivée d'un troisième ARNt qui porte le troisième acide aminé et ainsi de suite.
La phase de terminaison:
En face d'un codon non-sens, et comme il n'existe aucun ARNt capable de reconnaître ce codon, la synthèse s'arrête par la libération de la chaîne peptidique et le détachement des deux sous-unités du ribosome. Si l'acide aminé initiateur ne fait pas partie de la protéine synthétisée, il sera également libéré.
CONCLUSION:
Le phénomène de l'expression de l'information génétique est un phénomène rapide puisqu'on a constaté chez des bactéries l'association d'acides aminés pouvant atteindre 350 par minute. Il est également amplifié puisqu'un seul gène peut être transcrit plusieurs fois en même temps pour produire plusieurs ARNm correspondants, de même, un seul ARNm peut être lu par plusieurs ribosomes en même temps pour former ce qu'on appelle un polysome (chez l'Homme, un ARNm peut porter de 5 à 20 ribosomes successifs).
TRANSMISSION DE DEUX COUPLES D’ALLELES CHEZ LES DIPLOIDES :
Cas de gènes indépendants :
Exercice : on croise deux drosophiles de lignées pures différentes telles que l’une a les ailes longues et le corps gris et l’autre les ailes vestigiales (courtes) et le corps ébène (sombre). A la première génération, toutes les drosophiles ont les ailes longues et le corps gris. A la deuxième génération (F1XF1) on obtient parmi les descendants 9/16 aux ailes longues et à corps gris 3/16 aux ailes longues et à corps ébène 3/16 aux ailes vestigiales et à corps gris et 1/16 aux ailes vestigiales et à corps ébène.
Interprétation : dans cet exercice, on envisage la transmission de deux caractères héréditaires à savoir « la taille des ailes » et « la couleur du corps » donc il s’agit d’un cas de dihybridisme. La première génération est homogène et présente un phénotype parental [ailes longues et corps gris] donc il s’agit d’un cas de dominance pour les deux caractères. Les deux gènes sont indépendants. On pose les deux couples d’allèles suivants :
(L, l) / L= ailes longues et l= ailes vestigiales avec L > l
(G, g) / G= corps gris et g= corps ébène avec G > g

F1 : 100% [L G]
F1 X F1
Génotypes : L//l G//g pour chaque individu F1
Gamètes : L et G ; L et g ; l et G ; et l et g pour chaque individu F1.
comme chaque individu participe par quatre types de gamètes et comme la rencontre des gamètes est aléatoire, donc, le résultat sera une distibution au 1/16 qui donne selon la dominance:
9/16 [L G]
3/16 [L g]
3/16 [l G]
1/16 [l g]
Conclusion : la répartition de la F2 au seizième revient à la ségrégation indépendante des allèles au cours de la méiose et à leur réunion aléatoire au cours de la fécondation.
Le test-cross : le test-cross est un test qui permet se définir un génotype certain parmi plusieurs génotypes possibles qui se manifestent tous sous forme d’un seul phénotype. L’individu testé sera, alors croisé avec un testeur qui doit obligatoirement être homozygote récessif pour qu’il participe par un seul type de gamète porteur de l’allèle récessif pour laisser se manifester tous les allèles provenant du testé. Par exemple, si on cherche à connaître le génotype d’un individu pris au hasard de la population de la F2 qui présente le phénotype dominant ( ), ce test donnera un résultat parmi quatre possibles :
1er cas : testé [l G] X testeur [l g] donne 100% [L G] donc le testé a participé par un seul type de gamètes d’où il est certainement homozygote dominant pour les deux couples d’allèles.
2ème cas : le résultat est de 50% [L G], 50% [L g] donc le testé a participé par deux types de gamètes qui se distinguent par « la couleur du corps » d’où pour ce caractère le testé est hétérozygote et il est homozygote dominant pour l’autre caractère.
3ème cas : le résultat est de 50% [L G], 50% [l G] c’est le même raisonnement que pour le deuxième cas.
4ème cas : le résultat est de 25% [L G], 25% [L g], 25% [l G], 25% [l g] donc le testé a participé par quatre types de gamètes d’où il est hétérozygote pour les deux caractères.
Cas de gènes liés = linkage.
Exercice : on croise des drosophiles de lignées pures telles que les mâles sauvages ont les ailes longues et les yeux rouges et les femelles mutées ont les ailes vestigiales et les yeux bruns. La première génération est toute sauvage. Le croisement des mâles F1 avec des femelles mutées a donné 495 drosophiles du type sauvage et 508 drosophiles du type muté. Le croisement des femelles F1 avec des mâles mutés a donné : 712 drosophiles du type sauvage, 698 drosophiles du type muté, 300drosophiles aux ailes longues et aux yeux bruns et 298 drosophiles aux ailes vestigiales et aux yeux rouges.
Interprétation : dans cet exercice, on s’intéresse à l’étude de deux caractères héréditaires à savoir « la taille des ailes » et « la couleur des yeux » donc il s’agit d’un cas de dihybridisme. La première génération est homogène et présente un phénotype parental donc il s’agit d’un cas de dominance tel que les ailes longues dominent les ailes vestigiales et les yeux rouges dominent les yeux bruns. On pose alors les couples d’allèles suivants : (L, l) / L= longues et l= vestigiales avec L > l et (R, r) / R= rouges et r= bruns avec R > r
1er croisement :

F1 [L R] LR //lr
2ème croisement :

Cette descendance est expérimentalement formée de 1003 individu s( 495 + 508) dont la moitié théorique est 501,5 ce qui est à peu près égal à 495 et à 508; donc, le résultat théorique confirme bien le résultat expérimental et les deux gènes activés sont liés puisque le croisement envisagé est un test cross où le testé est hétérozygote pour les deux couples d’allèles (F1) donc si les deux gènes étaient indépendants, cet individu devrait produire quatre types de gamètes aux mêmes proportions.
3ème croisement :
Femelles F1 X mâles mutés
Comme le deuxième croisement, il s’agit d’un test cross qui devrait, normalement, donner le même résultat. Cependant, le résultat de ce croisement présente quatre phénotypes en des proportions différentes comme l'indique le tableau suivant :

Des résultats, on constate que les gamètes qui donnent les phénotypes parentaux ([L R] et [l r]) sont plus abondants (70%) que ceux qui ont donnés les phénotypes nouvellement construits (recombinés) (30%). Ce résultat laisse dire qu’au cours de la méiose, un accident s’est réalisé pour donner les gamètes recombinés. Il s’agit d’un crossing-over (brassage intra-chromosomique) qui a affecté 30% des gamètes de la femelle. Ainsi, on peut réaliser l’échiquier suivant :

Pour dire que dans un cas de dihybridisme il y a une liaison génétique, on réalise soit un test-cross au niveau duquel le testé participe par quatre types de gamètes en des proportions différentes deux à deux telles que des parentaux très fréquents et des recombinés moins fréquents. Soit une F2 qui se présente également avec quatre phénotypes mais à des proportions différentes de la répartition indépendante (9. 3. 3. 1) et qui présente également des recombinés peu fréquents et à la même proportion et des parentaux plus fréquents et à des proportions différentes.
La carte factorielle = carte génétique : il s’agit d’imaginer, selon les résultats statistiques, l’emplacement des deux gènes sur leur chromosome. La distance séparant ces deux gènes sera égale au pourcentage de recombinaison ( X 100). Ce résultat est donné en % ou en centimorgan
(C M) tel que 1 C M = 1%.
GENETIQUE HUMAINE
INTRODUCTION :
Appliquée à l'espèce humaine, la génétique conserve toutes ses normes. Cependant, confronté à des difficultés expérimentales, le dépistage des mouvements alléliques a imposé des nouvelles méthodes et a additionné de nouveaux critères de raisonnement.
les difficultés expérimentales :
un nombre important de chromosomes (2n = 46).
Les générations successives sont très espacées (en moyenne 25 ans).
Le nombre des descendants est statistiquement faible (la descendance d'une porté dépasse rarement un descendant).
Avec chaque acte reproducteur, les gamètes répondent à la même distribution aléatoire.
L'impossibilité de réaliser une reproduction orientée (entre frères et soeurs).
les méthodes utilisées :
la réalisation de pedigrees (arbres généalogiques).
la réalisation de caryotypes.
l'étude de l'activité des chromosomes (gènes ou protéines) par électrophorèse.
les nouveaux critères de raisonnement :
Si avec la génétique classique, l'énoncé vous donne le résultat et vous demande de l'expliquer en le confrontant à une statistique connue, en génétique humaine et comme souvent le résultat est confus, vous devez donner plus d'importance à l'observation pour éliminer logiquement les cas impossibles et tester ceux qui sont possibles pour pouvoir atteindre la certitude.
ETUDE DE CARUOTYPE NORMAL.
L'observation d'un caryotype normal permet d'envisager le nombre des chromosomes. Dans ce cas on distingue 46 chromosomes classés par paire selon leurs tailles, on peut également distinguer le sexe chromosomiques de l'individu concerné qui est marqué XX si l'individu est de sexe féminin, donc, la formule chromosomique s'écrit 2n = 44 autosomes + XX ou XY si l'individu est de sexe masculin, donc, la formule chromosomique s'écrit 2n = 44 autosomes + XY.
 ETUDE DE PEDIGREE :
ETUDE DE PEDIGREE :
Pour la génétique humaine, on traite seulement le cas de monohybridisme. Dans ce cas, il s'agit donc, de la transmission d'un couple d'allèles qui est le plus souvent dominant ou récessif et qui est soit porté par un autosome ou par un chromosome sexuel X ou Y. l'observation d'un pedigree permet d'éliminer des cas certains et pour les cas incertains il faut les mettre sous forme d'hypothèses à vérifier avec un traitement génétique. L'élimination d'un cas par simple observation, doit obéir à des normes logiques tel que dans un pedigree, quand les deux parents sont phénotypiquement sains et donnent un ou plusieurs descendants (garçons ou filles) malades donc l'allèle responsable de la maladie est récessif. Egalement, quand les deux parents sont phénotypiquement sains et donnent une fille malade donc la maladie est autosomale.
1.cas de maladies autosomales récessives : (phénylcétonurie)
 2.cas de maladie autosomale dominante : (maladie de Charcot-Marie)
2.cas de maladie autosomale dominante : (maladie de Charcot-Marie)
La maladie est le plus probable dominante quand elle apparaît dans toutes les générations. Dans ce cas, un père malade (comme II3)et une mère saine (comme II4) doivent donnés toutes leur filles malades si la maladie est portée par X. Donc, donc la maladie est autosomale dominante. On pose, alors, le couple d'allèles suivant (A, a) / A = malade et a = sain avec A > a. Ainsi, tout individu sain doit être homozygote sain et tout individu malade doit être hétérozygote.
3.cas de maladies récessives liées à X : (myopathie de Duchenne)
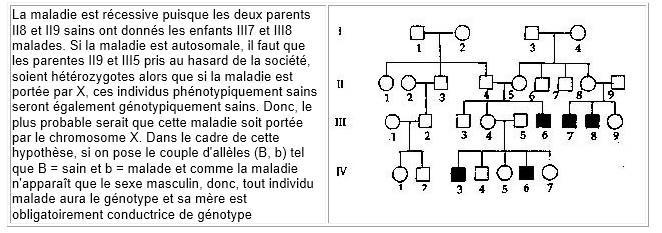
La maladie est récessive puisque les deux parents II8 et II9 sains ont donnés les enfants III7 et III8 malades. Si la maladie est autosomale, il faut que les parentes II9 et III5 pris au hasard de la société, soient hétérozygotes alors que si la maladie est portée par X, ces individus phénotypiquement sains seront également génotypiquement sains. Donc, le plus probable serait que cette maladie soit portée par le chromosome X. Dans le cadre de cette hypothèse, si on pose le couple d'allèles (B, b) tel que B = sain et b = malade et comme la maladie n'apparaît que le sexe masculin, donc, tout individu malade aura le génotype et sa mère est obligatoirement conductrice de génotype
 4.cas de maladie dominante liée à X : (rachitisme vitamino-résistant)
4.cas de maladie dominante liée à X : (rachitisme vitamino-résistant)
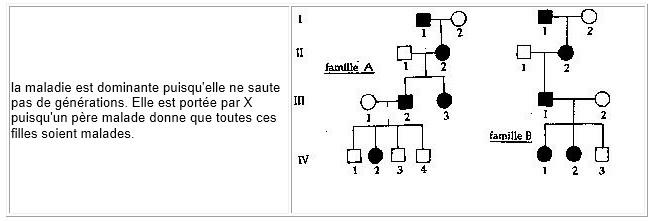
la maladie est dominante puisqu'elle ne saute pas de générations. Elle est portée par X puisqu'un père malade donne que toutes ces filles soient malades.
 5.cas de maladies liées à Y : (hypertrichose des oreilles)
5.cas de maladies liées à Y : (hypertrichose des oreilles)
comme le chromosome Y n'existe que chez le sexe masculin, quand il définit une maladie, elle sera transmise de père aux fils tel qu'un père malade donne que tous ces garçons soient malades et réciproquement.
 6.la consanguinité :
6.la consanguinité :
On appelle mariage consanguin, le mariage entre individus d'une même famille. Ce type de mariage présente un danger sur les descendants quand il y a une maladie récessive dans cette famille. Donc, le mariage consanguin a comme risque d'augmenter la fréquence de l'apparition des maladies récessives.
 LE DIAGNOSTIC PRENATAL:
LE DIAGNOSTIC PRENATAL:
C'est un test qui est réalisé sur des fœtus quand ils sont dans une famille à risque. Ce test consiste à prélever des cellules du fœtus afin de réaliser un caryotype ou de chercher une anomalie génique.
les techniques du diagnostic prénatal :
pour répondre aux objectifs de ce test, il faut prélever des cellules fœtales soit à partir du sang fœtal en aspirant une petite quantité d'une veine du cordon ombilicale (ce prélèvement devient sans danger quand il est fait au delà de la 20ème semaine de grossesse). Soit en aspirant une quantité du liquide amniotique (amniocentèse) qui baigne le fœtus (Ce prélèvement est normalement sans risque quand il est fait au delà de la 17ème semaine de grossesse). Soit en prélevant des villosités choriales (la biopsie du trophoblaste). Ce prélèvement est sans danger quand il est fait au delà de la 10ème semaine. Le premier type de prélèvement est souvent utilisé pour réaliser des caryotypes et les deux autres pour réaliser des tests sur l'ADN.
les résultats du diagnostic prénatal :
La réalisation du caryotype permet de rechercher le sexe du fœtus et le nombre de ses chromosomes. En effet, le sexe est masculin quand il y a XY et il est féminin quand il y a XX.
Pour le nombre des chromosomes, le caryotype est dit normal quand il y a 46 chromosomes. Mais, un caryotype peut être surnuméraire quand il y a 47 chromosomes. Cela peut être observé avec des autosomes tel que la trisomie 21 ou pour les chromosomes sexuels tel que pour le syndrome de Klinefelter où le caryotype montre 44 autosomes + XXY. Un caryotype peut également être réduit à 45 chromosomes tel que le syndrome de Turner où la formule chromosomique se note 44 autosomes + X.
L'obtention d'un caryotype anormal revient à un accident au cours de la méiose soit au cours de la première division quand les chromosomes homologues d'un bivalent migrent vers le même pôle, soit au cours de la deuxième division quand les deux chromatides d'un même chromosome migrent vers le même pôle. Ce phénomène affecte spécialement le gamète féminin.
Pour rechercher les anomalies géniques, on peut soit utiliser la sonde moléculaire radioactive pour chercher les allèles normaux ou mutés soit utiliser la méthode d'électrophorèse qui traduit la migration de l'ADN ou à la protéine correspondant à l'expression de cet ADN dans un champ électrique.
